|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
РОЛЬ ПЕЧЕНИ В УГЛЕВОДНОМ ОБМЕНЕ
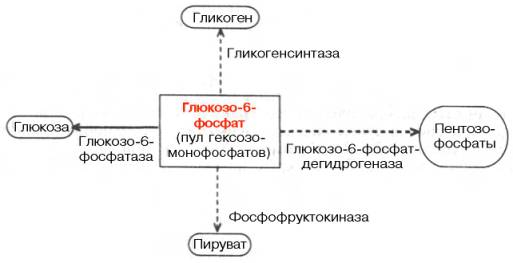
Печень играет центральную роль в многочисленных реакциях промежуточного обмена углеводов. Среди них особенно важны: · превращение галактозы в глюкозу; · превращение фруктозы в глюкозу; · синтез и распад гликогена; · глюконеогенез; · окисление глюкозы; · образование глюкуроновой кислоты. Превращение галактозы в глюкозу. Галактоза поступает в организм в составе молочного сахара. В печени происходит ее превращение через уридиндифосфогалактозу в глюкозо-1-фосфат. Галактоза в печени сначала фосфорилируется при участии АТФ и фермента галактокиназы с образованием галактозо-1-фосфата. Большая часть галактозо-1-фосфата в печени превращается в ходе реакции, катализируемой гексозо-1-фосфат-уридилилтрансферазой: УДФ-глюкоза + Галактозо-1-фосфат –> УДФ-галактоза + Глюкозо-1-фосфат. Это уникальная трансферазная реакция возвращения галактозы в основное русло углеводного метаболизма. Наследственная утрата гексозо-1-фосфат-уридилилтрансферазы приводит к галактоземии – заболеванию, для которого характерны умственная отсталость и катаракта хрусталика. В этом случае печень новорожденных теряет способность метаболизировать D-галактозу, входящую в состав лактозы молока. При нарушении функции печени способность организма использовать галактозу снижается, на этом основана функциональная проба печени с нагрузкой галактозой. Превращение фруктозы в глюкозу. Печень превращает фруктозу во фруктозо-1-фосфат (Ф-1-Ф) с помощью содержащейся в ней специфической фруктокиназы при участии АТФ. Фруктозо-1-фосфат расщепляется в печени альдолазой типа В, как и фруктозо-1, 6-дифосфат – промежуточный продукт обмена глюкозы, превращаясь в диоксиацетонфосфат и 3-фосфоглицериновый альдегид. Часть фруктозы под действием гексокиназы превращается в фруктозо-6-фосфат, промежуточный продукт основного пути распада глюкозы. Под действием глюкозофосфатизомеразы фруктозо-6-фосфат превращается в глюкозо-6-фосфат (Г-6-Ф). Исследование утилизации фруктозы положено в основу одной из функциональных проб печени, которая в настоящее время в клинике используется мало. Синтез и распад гликогена. Основная роль печени в углеводном обмене заключается в обеспечении постоянства концентрации глюкозы в крови. Это достигается регуляцией между синтезом и распадом гликогена, депонируемого в печени. Поддержание постоянного уровня гликемии относится к жизненно важным функциям печени. Особенно опасно для организма падение этого уровня (гипогликемия), поскольку некоторые ткани, например головной мозг, не способны запасать глюкозу, являющуюся важнейшим дыхательным субстратом. В печени все гексозы (шестиуглеродные сахара), включая фруктозу и галактозу, превращаются в глюкозу, которая в результате конденсации образует нерастворимую форму — полисахарид гликоген. После приема пищи богатой углеводами содержание гликогена может составлять до 8% веса печени. В среднем, запасы гликогена составляют около 5% веса печени, что у взрослого человека эквивалентно примерно 90 г глюкозы. Процесс превращения глюкозы в гликоген называется гликогенезом и стимулируется инсулином. Этот гормон вырабатывается поджелудочной железой в ответ на высокий уровень сахара в крови. Гликоген — полисахарид, включающий остатки d-глюкозы. Гликоген важен также как резервуар химической энергии. Синтез гликогена из глюкозы обеспечивает в норме временный резерв углеводов, необходимый для поддержания концентрации глюкозы в крови в тех случаях, если ее содержание значительно уменьшается (например, у человека это происходит при недостаточном поступлении углеводов с пищей или в период ночного «голодания»). Расщепляясь до глюкозы, гликоген препятствует падению уровня глюкозы в крови. Процесс расщепления гликогена называется гликогенолизом и включает активацию фермента фосфорилазы гормоном глюкагоном. Глюкагон тоже вырабатывается поджелудочной железой и выделяется в ответ на недостаток сахара в крови. В момент опасности, при стрессе или в условиях холода фосфорилазу активируют также адреналин, выделяемый мозговым веществом надпочечников, и норадреналином, высвобождаемый также мозговым веществом надпочечников и окончаниями симпатических нейронов. Глюконеогенез. Когда в связи с расходованием глюкозы запасы гликогена в печени истощаются, глюкоза может синтезироваться из любого неуглеводного предшественника. Этот процесс называется глюконеогенезом. Происходит он при истощении запасов гликогена в печени. Основными субстратами глюконеогенеза служат лактат, глицерин и аминокислоты. Принято считать, что почти все аминокислоты, за исключением лейцина, могут пополнять пул предшественников глюконеогенеза. Низкий уровень глюкозы в крови (гипогликемия) стимулирует посредством симпатической нервной системы выброс адреналина, который, как уже отмечалось, способствует сиюминутному удовлетворению потребностей организма в глюкозе. Низкий уровень глюкозы в крови приводит также к стимуляции гипоталамуса, который выделяет кортиколиберин, вызывающий секрецию адренокортикотропного гормона (АКТГ) передней долей гипофиза. Под действием АКТГ усиливается синтез и высвобождение глюкокортикоидных гормонов (в основном кортизола, известного также как гидрокортизон). Эти гормоны стимулируют переход из тканей в кровь аминокислот, глицерола и жирных кислот, а также синтез в печени ферментов, катализирующих превращение аминокислоты и глицерола в глюкозу, т. е. осуществляют глюконеогенез. Жирные кислоты расщепляются с образованием ацетилкофермента А, а затем окисляются в цикле Кребса. Центральную роль в превращениях глюкозы и саморегуляции углеводного обмена в печени играет глюкозо-6-фосфат. Он резко тормозит фосфоролитическое расщепление гликогена, активирует ферментативный перенос глюкозы с уридиндифосфоглюкозы на молекулу синтезирующегося гликогена, является субстратом для дальнейших гликолитических превращений, а также окисления глюкозы, в том числе по пентозофосфатному пути. Наконец, расщепление глюкозо-6-фосфата фосфатазой обеспечивает поступление в кровь свободной глюкозы, доставляемой током крови во все органы и ткани (рис. 14).
Рис. 14. Участие глюкозо-6-фосфата в метаболизме углеводов Функции накопления (синтеза) глюкозы в виде гликогена и его распада должны быть взаимосогласованы. Таким образом, совершенно невозможно одновременное протекание гликолиза и глюконеогенеза, как и синтеза и деградации гликогена. Согласование процессов обеспечивается тем, что синтез (анаболизм) и распад (катаболизм) катализируются двумя различнымиферментами и контролируются независимо. Окисление глюкозы. Расщепление глюкозы дает организму большое количество энергии. Так, окисление ее до конечных продуктов — воды и углекислого газа — сопровождается выделением 686 ккал/моль, при этом половина энергии аккумулируется АТФ и другими макроэргическими соединениями. Распад глюкозы, происходит и в анаэробных условиях (гликолиз), что очень важно для жизнедеятельности многих тканей. Энергии при этом высвобождается значительно меньше, и образуется молочная кислота. Это дополнительный путь обмена веществ в печени. Образование глюкуроновой кислоты. С обменом углеводов связан синтез глюкуроновой кислоты, необходимой для конъюгации плохо растворимых веществ (фенолы, билирубин и др.) и образования смешанных полисахаридов (гиалуроновая кислота, гепарин и др.). Также глюкуроновая кислота, присоединяясь при помощи фермента глюкуронилтрансферазы, образует водорастворимое соединения в виде билирубина диглюкуронида и билирубин моноглюкуронида. Таким образом, билирубин лишается от токсических свойств и выделяется желчью в кишечник. При уменьшении образования глюкуроновой кислоты, страдает антитоксическая функция печени. Контрольные вопросы 1. Какие реакции промежуточного обмена углеводов происходят в печени? 2. Как происходит в печени превращение галактозы в глюкозу? 3. Как происходит в печени превращениефруктозы в глюкозу? 4. Охарактеризуйте процессы синтеза и распада гликогена в печени. 5. Что такое глюконеогенез? 6. Какой фермент играет центральную роль в превращениях глюкозы и саморегуляции углеводного обмена в печени? 7. Какой побочный продукт образуется в процессе превращения глюкозы в печени? РОЛЬ ПЕЧЕНИ В ОБМЕНЕ БЕЛКОВ
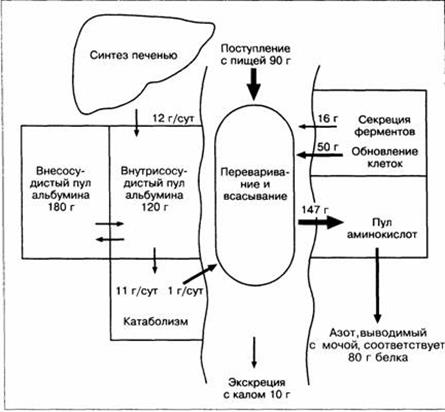
Печень играет ключевую роль в обмене белков. В этом направлении она выполняет следующие основные функции: · синтез специфических белков плазмы; · образование мочевины и мочевой кислоты; · синтез холина и креатина; · трансаминирование и дезаминирование аминокислот, что весьма важно для взаимных превращений аминокислот, а также для процесса глюконеогенеза и образования кетоновых тел. Синтез специфических белков плазмы. Все альбумины плазмы, 75 — 90% α -глобулинов и 50% β -глобулинов синтезируются гепатоцитами. Лишь γ -глобулины продуцируются не гепатоцитами, а звездчатыми ретикулоэндотелиоцитами печени (клетками Купфера). Следует, однако, подчеркнуть, что в большинстве своём γ -глобулины образуются вообще вне печени. Строительным материалом для этих целей служат аминокислоты, поступающие извне, а также появляющиеся в процессе катаболизма тканевых белков, обмена жирных кислот и углеводов. Ежедневно образуется около 12 г альбумина. В зависимости от потребностей организма альбумин синтезируется в 10-60% гепатоцитов. Около 60% альбумина покидает сосудистое русло, однако оставшиеся 40% составляют наибольшую фракцию белков плазмы. Альбумин играет важную роль в поддержании онкотического давления крови. Кроме того, он необходим для связывания и транспортировки многих веществ, в том числе некоторых гормонов, жирных кислот, микроэлементов, триптофана, билирубина, многих эндогенных и экзогенных органических анионов. Однако при редком врожденном нарушении – анальбуминемии не возникает тяжелых физиологических изменений, кроме избыточного накопления жидкости в тканях.
Рис. 15. Метаболизм альбумина плазмы у взрослого человека с массой тела 70 кг с нейтральным азотистым балансом. Общий пул альбумина, составляющий около 300 г, распределён между внутрисосудистым и внесосудистым пространством в соотношении приблизительно 2: 3. На этой упрошенной схеме баланс представлен в граммах белка (=6, 25 • г азота). В схему не включены малосущественные потери белка (например, 2 г/сут через кожные покровы) Синтез альбумина зависит от поступления предшественников аминокислот, особенно триптофана – самой редкой из незаменимых аминокислот. У больных с крупным карциноидом синтез альбумина может резко понизиться, так как клетки опухоли используют триптофан для синтеза серотонина. При понижении онкотического давления плазмы синтез альбумина увеличивается. Наконец, на метаболизм белков в печени влияют такие гормоны, как глюкагон и инсулин. В печени образуются и другие секретируемые белки. К клинически значимым секретируемым гликопротеидам относятся: церулоплазмин, альфа1-антитрипсин и большинство других альфа-глобулинов и бета-глобулинов. Среди гликопротеидов в последнее время привлекает внимание фибронектин. Он синтезируется в основном печенью. В этом процессе участвуют звездчатые ретикулоэндотелиоциты. Фибронектин — компонент соединительной ткани, выполняет структурные функции, он расходуется в процессе выделения осколков гепатоцитов и других клеток. Недостаток фибронектина может способствовать микроэмболической обструкции легких и нарушению системной микроциркуляции. Протеинообразовательная функция печени играет важную роль в обеспечении гемостаза. В печени синтезируются многие факторы свертывания: фибриноген (фактор I), протромбин (фактор II), фактор V, фактор VII, фактор IХ, фактор X, фактор XI, фактор XI, фактор XIII, а также ингибиторы свертывания и фибринолиза. Синтез протромбина и факторов VII, IХ и X зависит от наличия витамина К и, следовательно, от всасывания жиров в кишечнике (витамин К жирорастворим). Витамин К активирует ферменты эндоплазматического ретикулума гепатоцитов, катализирующие гамма-карбоксилирование остатков глутаминовой кислоты в предшественниках факторов свертывания. Благодаря гамма-карбоксилированию, в частности, возрастает способность протромбина связывать ионы кальция и фосфолипиды и быстро превращаться в тромбин в присутствии факторов V и X. Метаболическая функция печени имеет большое значение в регуляции гемостаза. Тяжелое поражение печени ведет к снижению синтеза протромбина. Гипопротромбинемия может усилиться из-за нарушения всасывания витамина К при истощении, введении антибиотиков широкого спектра действия или нарушении всасывания жиров из-за понижения концентрации желчных кислот в кишечнике (например, при холестазе ). В таких случаях для нормализации уровня протромбина назначаются препараты витамина К в/м или в/в. Однако если коагулопатия возникает в результате нарушения функции гепатоцитов и не связана с холестазом или нарушением всасывания, то введение препаратов витамина К не влияет на синтез протромбина. При болезнях печени может нарушиться синтез и других факторов свертывания. Так, тяжелое поражение печени иногда приводит к снижению концентрации в плазме фактора V. Концентрация фибриногена обычно почти не изменяется, кроме тех случаев, когда развивается ДВС-синдром. По неизвестным причинам поврежденная печень может синтезировать повышенное количество фибриноген, а также других белков, которые называют белками острой фазы воспаления (С-реактивный белок, гаптоглобин, церулоплазмин и трансферрин). Последний образуется как при повреждении печени, так и при системных заболеваниях - злокачественных новообразованиях, ревматоидном артрите, бактериальных инфекциях, ожогах, инфаркте миокарда. Видимо, синтез белков острой фазы воспаления стимулируется цитокинами, включая ИЛ-1 и ИЛ-6. Хотя поврежденная печень может синтезировать нормальное или повышенное количество фибриногена, но его молекулярная структура может быть значительно изменена из-за тонких нарушений синтеза белков. Возможно, это один из механизмов нарушений гемостаза, часто возникающих при хронических болезнях печени. Образование мочевины и мочевой кислоты. При дезаминировании азотистых соединений, в первую очередь аминокислот, образуется аммиак. При разрушении 100 г белка образуется около 20 г аммиака. Под аммиаком подразумевается как неионизированный NH3, так и ионизированный NH4. Большие концентрации аммиака сыворотки крови и тканей обладают высокотоксическими свойствами, тогда как к нормальным концентрациям NH3 организм адаптирован. Аммиак — один из видов сырья для синтеза мочевины. Различают две основные причины гипераммониемии — избыточное поступление NH3 из кишечника и уменьшение преобразования аммиака в печени. В основном гипераммониемия наблюдается при заболеваниях печени, особенно тяжелых (цирроз печени, синдром Рейя и др.). Несравненно реже наблюдаются врожденные дефекты ферментных систем, преобразующих аммиак (дефекты дегидрогеназы лизина, метилмалонилмутазы и др.). Высокая концентрация аммиака в головном мозге при печеночной коме ведет к нарушению обмена энергии и деятельности Na+, K+-ATФазы, увеличению количества и размера астроцитов, а также концентрации токсических продуктов метаболизма аммиака. Кроме того, нарушается медиаторная передача - в частности, образуются " ложные" медиаторы, которые могут конкурировать с обычными медиаторами за рецепторы. Наконец, аммиак или другие продукты его метаболизма могут сами взаимодействовать с бензодиазепиновыми рецепторами, вызывая угнетение ЦНС. Образование мочевины происходит в основном в печени. Таким путем достигается превращение ядовитых осколков белковой молекулы (аминогрупп и др.) в практически нетоксичное вещество — мочевину. На синтез 1 моля мочевины расходуется 2 моля бикарбоната, и таким образом снижается рН. Синтез мочевины относится к одной из устойчивых функций печени. Поэтому снижение концентраций мочевины в сыворотке крови наблюдается нечасто. Эта закономерность касается суммарной продукции мочевины. Нарушение отдельных этапов ее синтеза может резко не нарушать суммарную концентрацию мочевины сыворотки крови, но приводить к увеличению концентраций токсичных продуктов, образующихся на отдельных этапах синтеза молекулы мочевины. Другим продуктом обезвреживания аммиака является глутамин. Синтез его осуществляется не только в печени. Отличие синтеза глутамина от синтеза мочевины заключается в том, что первый синтезируется и при низких концентрациях аммиака, тогда как вторая — только при достаточно высоких концентрациях аммиака. При низком рН происходит расщепление глутамина, при высоком рН — энергичный синтез мочевины. Оба процесса направлены на стабилизацию уровня рН в нормальных условиях. Большая часть мочевой кислоты также образуется в печени, где много фермента ксантиноксидазы, при участии которого оксипурины (гипоксантин и ксантин) превращаются в мочевую кислоту.Мочевая кислота обычно образуется как конечный продукт обмена пуриновых соединений. Наиболее стабильная гиперурикемия наблюдается при подагре. Печень участвует в обмене пуринов, и при ряде заболеваний печени, в первую очередь алкогольных, наблюдается гиперурикемия. Чаще она следует за острой интоксикацией алкоголем. В повышенной продукции мочевой кислоты велика роль индукции таких ферментов гепатоцитов, как ксантиноксидаза и глутатионредуктаза. Синтез холина и креатина. Нельзя забывать о роли печени и в синтезе креатина. Имеются два источника креатина в организме. Существует экзогенный креатин, т.е. креатин пищевых продуктов (мясо, печень и др.), и эндогенный креатин, синтезирующийся в тканях. Синтез креатина происходит в основном в печени, откуда он с током крови поступает в мышечную ткань. Здесь креатин, фосфорилируясь, превращается в креатинфосфат, а из последнего образуется креатинин. Необходимо подчеркнуть роль метионина, способного участвовать в процессах переметилирования, в результате которых синтезируется холин. При поражении печеночной паренхимы количество АТФ в печеночных клетках резко снижается, и процессы переметилирования нарушаются Трансаминирование и дезаминирование аминокислот. Организм не может запасать аминокислоты, и те из них, которые не используются сразу для биосинтеза белка, подвергаются дезаминированию в печени. Этот процесс состоит в ферментативном отщеплении аминогруппы и одновременном окислении остатка молекулы с образованием углевода, который используется в процессе дыхания. Аминогруппа отщепляется вместе с атомом водорода, поэтому азотистым продуктом дезаминирования оказывается аммиак, который может быть использован для синтеза определенных аминокислот или оснований типа аденина или гуанина, либо удален из организма. При поражениях печени нарушается процесс дезаминирования аминокислот, что приводит к увеличению их концентрации в крови и моче. Так, если в норме содержание азота аминокислот в сыворотке крови составляет примерно 2, 9-4, 3 ммоль/л, то при тяжелых заболеваниях печени (атрофические процессы) эта величина возрастает до 21 ммоль/л, что обусловливает выраженную аминоацидурию. К примеру, при острой атрофии печени содержание тирозина в суточной моче может достигать 2 г (при норме 0, 02 — 0, 05 г/сут). Кроме дезаминирования, аминокислоты подвергаются в печени переаминированию. Процесс переаминирования (трансаминирования) не является специфическим для печени, он происходит и в других органах, но в печени интенсивность этих ферментативных реакций весьма значительна. В крови повышение активности трансаминаз ( АЛТ - аланинаминотрансферазы, АСТ - аспартатаминотрансферазы) наблюдается при различных деструктивных изменениях, например при инфарктах миокарда и при гепатитах. Очень важна специфичность данной энзимодиагностики: с помощью этих ферментативных реакций можно с высокой степенью достоверности судить о поражении указанных органов. При некротических изменениях в сердечной мышце в крови резко повышается активность АСТ, в то время как при гепатите отмечается возрастание активности АЛТ. Причем весьма важным является то, что определение активности трансаминаз позволяет обнаружить патологию печени ещё в дожелтушный период. Контрольные вопросы 1. Какие специфические белки плазмы синтезируются в печени? 2. Какие факторы свертывания синтезируются в печени? 3. Как происходит в печени образование мочевины и мочевой кислоты? 4. Что такое трансаминирование и дезаминирование аминокислот?
Популярное:
|
Последнее изменение этой страницы: 2017-03-11; Просмотров: 1607; Нарушение авторского права страницы