|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Клеточные мембраны, их виды. Свойства мембран. Функции мембран.
Биологические мембраны – сложные надмолекулярные структуры, окружающие все живые клетки и образующие в них замкнутые, специализированные компартменты – органеллы. Мембрану, ограничивающую цитоплазму клетки снаружи, называют цитоплазматической или плазматической мембраной. Название внутриклеточных мембран обычно происходит от названия ограничиваемых или образуемых ими субклеточных структур. Различают: · ядерную, · митохондриальную, · лизосомальную мембраны, · мембраны комплекса Гольджи, · эндоплазматический ретикулум и другие. Мембрана - это тонкая структура толщиной 7 нм. По своему химическому составу мембрана содержит: · 25% белков, · 25% фосфолипидов, · 13% холестерин, · 4% липиды, · 3% углеводы. В структурном отношении основу мембраны составляет двойной слой фосфолипидов. Особенностью молекул фосфолипидов является то, что в своем составе они имеют гидрофильную и гидрофобную части. Гидрофильные части содержат полярные группы (фосфатные группы в фосфолипидах и гидроксидные в холестеринах). Гидрофильные части направлены к поверхности. А гидрофобные (жирные хвосты) направлены к центру мембраны. Молекула имеет два жирных хвоста, и эти углеводородные цепи могут находится в двух конфигурациях. Вытянутые - транс-конфигурация (цилиндр 0.48 нм). Второй вид - гош-транс-гош конфигурация. В этом случае два жирных хвоста расходятся и площадь увеличивается до 0.58 нм. Молекулы липидов в нормальных условиях имеют жидкокристаллическую форму. И в этом состоянии они обладают подвижностью. Причем они могут, как передвигаться внутри своего слоя, так и переворачиваться. При понижении температуры происходит переход из жидкого состояния мембраны в желеобразное, и это уменьшает подвижность молекулы. При движении молекулы липидов образуются микрополоски, которые называются кингами, в которые могут захватываться вещества. Липидный слой в мембране является барьером для водорастворимых веществ, но зато пропускает жирорастворимые вещества. Замкнутый липидный бислой определяет основные свойства мембран:
Рисунок 14.8. Влияние жирнокислотного состава фосфолипидов на текучесть липидного бислоя. 2) латеральная диффузия – свободное перемещение молекул относительно друг друга в плоскости мембран (рисунок 14.9, а).
Рисунок 14.9. Виды перемещений фосфолипидных молекул в липидном бислое. 3) ограниченная способность к поперечной диффузии (переходу молекул из наружного слоя во внутренний и наоборот, см. рисунок 14.9, б), что способствует сохранению асимметрии – структурно-функциональных различий наружного и внутреннего слоёв мембраны. 4) непроницаемость замкнутого бислоя для большинства водорастворимых молекул.
В составе мембраны кроме липидов имеются еще белковые молекулы. В основном это гликопротеины. Интегральные белки проходят через оба слоя. Другие белки частично погружены либо в наружный, либо во внутренний слой. Они носят название периферических белков.
Данная модель мембраны называется жидко-кристалической моделью. Функционально белковые молекулы выполняют структурную, транспортную, ферментативную функции. Кроме того, они образуют ионные каналы с диаметром от 0.35 до 0.8 нм в диаметре, через которые могут проходить ионы. Каналы имеют свою специализацию. Интегральные белки участвуют в активном транспорте и в облегченной диффузии. Периферическим белкам на внутренней стороне мембраны характерна ферментативная функция. На внутренней стороне - антигенная (антитела) и рецепторная функции.
Углеродные цепи могут присоединятся к белковым молекулам, и тогда образуются гликопротеинами. Или к липидам, тогда они называются гликолипидами. Основными функции клеточных мембран будут являться: 1. Барьерная функция (выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам.) 2. Перенос веществ.
Рисунок 14.10. Механизмы транспорта молекул через мембрану Простая диффузия - перенос веществ через мембрану без участия специальных механизмов. Транспорт происходит по градиенту концентрации без затраты энергии. Путём простой диффузии транспортируются малые биомолекулы – Н2О, СО2, О2, мочевина, гидрофобные низкомолекулярные вещества. Скорость простой диффузии пропорциональна градиенту концентрации. Облегчённая диффузия - перенос веществ через мембрану при помощи белковых каналов или специальных белков-переносчиков. Осуществляется по градиенту концентрации без затраты энергии. Транспортируются моносахариды, аминокислоты, нуклеотиды, глицерол, некоторые ионы. Характерна кинетика насыщения – при определённой (насыщающей) концентрации переносимого вещества в переносе принимают участие все молекулы переносчика и скорость транспорта достигает предельной величины. Активный транспорт – также требует участия специальных белков-переносчиков, но перенос происходит против градиента концентрации и поэтому требует затраты энергии. При помощи этого механизма через клеточную мембрану транспортируются ионы Na+, K+, Ca2+, Mg2+, через митохондриальную – протоны. Для активного транспорта веществ характерна кинетика насыщения. Наряду с транспортом органических веществ и ионов, осуществляемым переносчиками, в клетке существует совершенно особый механизм, предназначенный для поглощения клеткой и выведения из неё высокомолекулярных соединений при помощи изменения формы биомембраны. Такой механизм называют везикулярным транспортом.
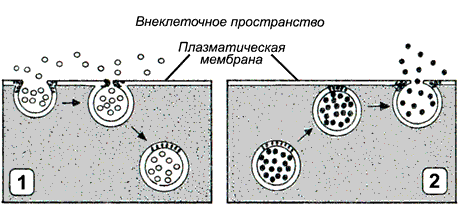
Рисунок 14.12. Типы везикулярного транспорта: 1 - эндоцитоз; 2 - экзоцитоз. При переносе макромолекул происходит последовательное образование и слияние окружённых мембраной пузырьков (везикул). По направлению транспорта и характеру переносимых веществ различают следующие типы везикулярного транспорта: Эндоцитоз (рисунок 14.12, 1) — перенос веществ в клетку. В зависимости от размера образующихся везикул различают: а) пиноцитоз — поглощение жидкости и растворённых макромолекул (белков, полисахаридов, нуклеиновых кислот) с помощью небольших пузырьков (150 нм в диаметре); б) фагоцитоз — поглощение крупных частиц, таких, как микроорганизмы или обломки клеток. В этом случае образуются крупные пузырьки, называемые фагосомами диаметром более 250 нм. Пиноцитоз характерен для большинства эукариотических клеток, в то время как крупные частицы поглощаются специализированными клетками - лейкоцитами и макрофагами. На первой стадии эндоцитоза вещества или частицы адсорбируются на поверхности мембраны, этот процесс происходит без затраты энергии. На следующей стадии мембрана с адсорбированным веществом углубляется в цитоплазму; образовавшиеся локальные впячивания плазматической мембраны отшнуровываются от поверхности клетки, образуя пузырьки, которые затем мигрируют внутрь клетки. Этот процесс связан системой микрофиламентов и является энергозависимым. Поступившие в клетку пузырьки и фагосомы могут сливаться с лизосомами. Содержащиеся в лизосомах ферменты расщепляют вещества, содержащиеся в пузырьках и фагосомах до низкомолекулярных продуктов (аминокислот, моносахаридов, нуклеотидов), которые транспортируются в цитозоль, где они могут быть использованы клеткой. Экзоцитоз (рисунок 14.12, 2) — перенос частиц и крупных соединений из клетки. Этот процесс, как и эндоцитоз, протекает с поглощением энергии. Основными разновидностями экзоцитоза являются: а) секреция - выведение из клетки водорастворимых соединений, которые используются или воздействуют на другие клетки организма. Может осуществляться как неспециализированными клетками, так и клетками эндокринных желёз, слизистой желудочно-кишечного тракта, приспособленными для секреции производимых ими веществ (гормонов, нейромедиаторов, проферментов) в зависимости от определённых потребностей организма. Секретируемые белки синтезируются на рибосомах, связанных с мембранами шероховатого эндоплазматического ретикулума. Затем эти белки транспортируются к аппарату Гольджи, где они модифицируются, концентрируются, сортируются, и затем упаковываются в пузырьки, которые отщепляются в цитозоль и в дальнейшем сливаются с плазматической мембраной, так что содержимое пузырьков оказывается вне клетки. В отличие от макромолекул, секретируемые частицы малых размеров, например, протоны, транспортируются из клетки при помощи механизмов облегчённой диффузии и активного транспорта. б) экскреция - удаление из клетки веществ, которые не могут быть использованы (например, удаление в ходе эритропоэза из ретикулоцитов сетчатой субстанции, представляющей собой агрегированные остатки органелл). Механизм экскреции, по-видимому, состоит в том, что вначале выделяемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазматической мембраной. 3. Метаболическая функция (благодаря наличию в них ферментных систем) 4. Мембраны участвуют в создании электрических потенциалов в состоянии покоя, а при возбуждении - токов действия. 5. Рецепторная функция. 6. Иммунологическая (связана с наличием антигенов и выработкой антител). 7. Обеспечивают межклеточное взаимодействие и контактное торможение. (При контакте однородных клеток возникает торможение деления клеток. Эта функция утрачивается у раковых клеток. Кроме того, раковые клетки вступают в контакт не только со своими, но и с другими клетками, заражая их.) Рецепторы, их классификация: по локализации (мембранные, ядерные), механизму развития процессов (ионо- и метаьотропные), по скорости приема сигнала (быстрые, медленные), по роду вопринимающих веществ. Рецепторы представляют собой конечные специализированные образования, предназначенные для трансформации энергии различных видов раздражителей в специфическую активность нервной системы. Классификация: по локализации · мембранные · ядерные по механизму развития процессов · ионотропные (представляют собой мембранные каналы, открываемые или закрываемые при связывании с лигандом. Возникающие при этом ионные токи вызывают изменения трансмембранной разности потенциалов и, вследствие этого, возбудимости клетки, а также меняют внутриклеточные концентрации ионов, что может вторично приводитъ к активации систем внутриклеточных посредников. Одним из наиболее полно изученных ионотропных рецепторов является н-холинорецептор.) · метаботропные (связаны с системами внутриклеточных посредников. Изменения их конформации при связывании с лигандом приводит к запуску каскада биохимических реакций, и, в конечном счете, изменению функционального состояния клетки.) по скорости приема сигнала · быстрые · медленные по роду вопринимающих веществ · Хеморецепторы — воспринимают воздействие растворенных или летучих химических веществ. · Осморецепторы — воспринимают изменения осмотической концентрации жидкости (как правило, внутренней среды). · Механорецепторы — воспринимают механические стимулы (прикосновение, давление, растяжение, колебания воды или воздуха и т. п.) · Фоторецепторы — воспринимают видимый и ультрафиолетовый свет · Терморецепторы — воспринимают понижение (холодовые) или повышение (тепловые) температуры · Барорецепторы – воспринимают изменение давления
3. Ионотропные рецепторы, метаботпропные рецепторы и их разновидности. Системы вторичных посредников действия метаботропных рецепторов (цАМФ, цГМФ, инозитол-3-фосфат, диацилглицерол, ионы Са++). На постсинаптической мембране выделяют два типа рецепторов — ионотропные и метаботропные. Ионотропнный С внутренней стороны мембраны к такому рецептору присоединен целый ряд других белков, выполняющих ферментативные и частью передающие («посреднические») функции (рис.). Белки-посредники относятся к G-белкам. Под влиянием возбужденного рецептора G-белок воздействует на белок-фермент, обычно переводя его в «рабочее» состояние. В результате запускается химическая реакция: молекула-предшественник превращается в сигнальную молекулу — вторичный посредник.
Рис. Схема строения и функционирования метаботропного рецептора: 1 — медиатор; 2 — рецептор; 3 — ионный канал; 4— вторичный посредник; 5 — фермент; 6 — G-белок; → — направление передачи сигнала Вторичные посредники — это мелкие, способные к быстрому перемещению молекулы или ионы, передающие сигнал внутри клетки. Этим они отличаются от «первичных посредников» — медиаторов и гормонов, передающих информацию от клетки к клетке. Наиболее известным вторичным посредником является цАМФ (циклическая аденозинмонофосфорная кислота), образуемая из АТФ с помощью фермента аденилатциклазы. Похожа на него цГМФ (гуанозинмонофосфорная кислота). Другими важнейшими вторичными посредниками являются инозитолтрифосфат и диацилглицерол, образуемые из компонентов клеточной мембраны под действием фермента фосфолипазы С. Чрезвычайно велика роль Ca2+, входящего в клетку снаружи через ионные каналы или высвобождающегося из особых мест хранения внутри клетки («депо» кальция). В последнее время много внимания уделяется вторичному посреднику NO (оксиду азота), который способен передавать сигнал не только внутри клетки, но и между клетками, легко преодолевая мембрану, в том числе от постсинаптического нейрона к пресинаптическому. Заключительный шаг в проведении химического сигнала — воздействие вторичного посредника на хемочувствительный ионный канал. Это воздействие протекает либо непосредственно, либо через дополнительные промежуточные звенья (ферменты). В любом случае происходит открытие ионного канала и развитие ВПСП либо ТПСП. Продолжительность и амплитуда их первой фазы будет определяться количеством вторичного посредника, которое зависит от количества выделившегося медиатора и длительности его взаимодействия с рецептором. Таким образом, механизм передачи нервного стимула, используемый метаботропными рецепторами, включает в себя несколько последовательных этапов. На каждом из них возможна регуляция (ослабление либо усиление) сигнала, что делает реакцию постсинаптической клетки более гибкой и адаптированной к текущим условиям. Вместе с тем это же приводит к замедлению процесса передачи информации
Система цАМФ
Фосфолипаза С
Популярное:
|
Последнее изменение этой страницы: 2016-04-09; Просмотров: 10413; Нарушение авторского права страницы
 1) текучесть – зависит от соотношения насыщенных и ненасыщенных жирных кислот в составе мембранных липидов. Гидрофобные цепочки насыщенных жирных кислот ориентированы параллельно друг другу и образуют жёсткую кристаллическую структуру (рисунок 14.8, а). Ненасыщенные жирные кислоты, имеющие изогнутую углеводородную цепь, нарушают компактность упаковки и придают мембране бó льшую жидкостность (рисунок 14.8, б). Холестерол, встраиваясь между жирными кислотами, уплотняет их и повышает жёсткость мембран.
1) текучесть – зависит от соотношения насыщенных и ненасыщенных жирных кислот в составе мембранных липидов. Гидрофобные цепочки насыщенных жирных кислот ориентированы параллельно друг другу и образуют жёсткую кристаллическую структуру (рисунок 14.8, а). Ненасыщенные жирные кислоты, имеющие изогнутую углеводородную цепь, нарушают компактность упаковки и придают мембране бó льшую жидкостность (рисунок 14.8, б). Холестерол, встраиваясь между жирными кислотами, уплотняет их и повышает жёсткость мембран.